


Mettiamoci l’animo in pace.
La vite vive su questo pianeta da molto tempo prima dell’uomo.
L’incontro tra le due specie ha avuto un impatto fondamentale per l’evoluzione tecnica e culturale di Homo sapiens. Per Vitis vinifera invece il percorso comune non è stato privo di rischi, come dimostrano la crisi fillosserica e le complicazioni create dall’arrivo in Europa di patologie come peronospora e oidio.
Squilibri di cui occorre tener conto quando si pretende di fermare il tempo e la storia evolutiva di questa specie propagando per via agamica sempre le stesse varietà per secoli.
Anche perché la coltivazione è in fondo solo una breve parentesi della sua storia biologica: meno di una settimana, dal primo addomesticamento fino all’ultima vendemmia, se ridotta in termini di vita media umana.
Articolo tratto dal numero 6/2021 di VVQ
Abbonati e accedi all’edicola digitale
28 milioni di anni di storia
Il genere Vitis è apparso infatti attorno a 28 milioni di anni fa (l’Homo sapiens esiste solo da 2,5 milioni di anni). «Circa 18 milioni di anni fa è avvenuta la separazione tra i generi Vitis e Muscadinia. Le diverse specie hanno iniziato a differenziarsi 6 milioni di anni dopo con un processo durato fino a 1,3 milioni di anni fa».
«5-6 milioni di anni fa la specie europee si sono separate da quelle asiatiche, da cui hanno avuto origine. La speciazione è probabilmente ancora in corso».
Lo ha ricordato nella sua lectio magistralis nel corso dell’ultima edizione di Conavi di Udine Raffaele Testolin. Una relazione su invito degli organizzatori dell’8° Congresso nazionale di viticoltura che è stata il modo migliore per celebrare l’opera di un docente e ricercatore di genetica che ha dato un contributo decisivo allo sviluppo della vite del futuro.
Leggi anche
Ottavo Conavi al via: genetica, biodiversità e sensoristica
Prospettive future
- Recupero e valutazione di materiale caucasico e dell’Asia centrale;
- analisi dei funding clones considerando l’intero germoplasma euro-asiatico e loro utilizzo nei programmi di breeding;
- recupero di varietà particolarmente adattate ad ambienti e climi estremi;
- introgressione di resistenze da specie selvatiche ma anche da V. vinifera nelle varietà coltivate in ambienti temperato freddi;
- fine tuning delle varietà coltivate per migliorare le caratteristiche agronomiche (resistenze a patogeni, vigoria …) ed enologiche (colore dei vini, acidità, contenuto e qualità dei tannini …) attraverso tecniche di genome editing.
“Padrino” della vite del futuro

Testolin è stato infatti co-fondatore nel 2006 e presidente per 10 anni dell’Istituto di Genomica applicata di Udine, istituto di ricerca non-profit che gestisce una piattaforma nazionale di sequenziamento di DNA e che ha partecipato al progetto italo-francese che ha portato al primo sequenziamento del genoma della vite.
Nel 2020 ha pubblicato la sequenza completa del primo gene di resistenza a peronospora rinvenuto in V. vinifera (rpv3).
A partire dal 1998 ha sviluppato con alcuni colleghi un programma di miglioramento genetico della vite che ha portato al licenziamento (con protezione europea, estesa ad altri Paesi extra-UE) di 14 varietà di vite da vino con resistenze a peronospora e oidio.
Al Conavi Testolin ha spiegato come le più recenti acquisizioni nell’ambito delle scienze genomiche stiano consentendo di ampliare le conoscenze su molti aspetti determinanti relativi all’identità di V. vinifera.
L’origine in Occidente
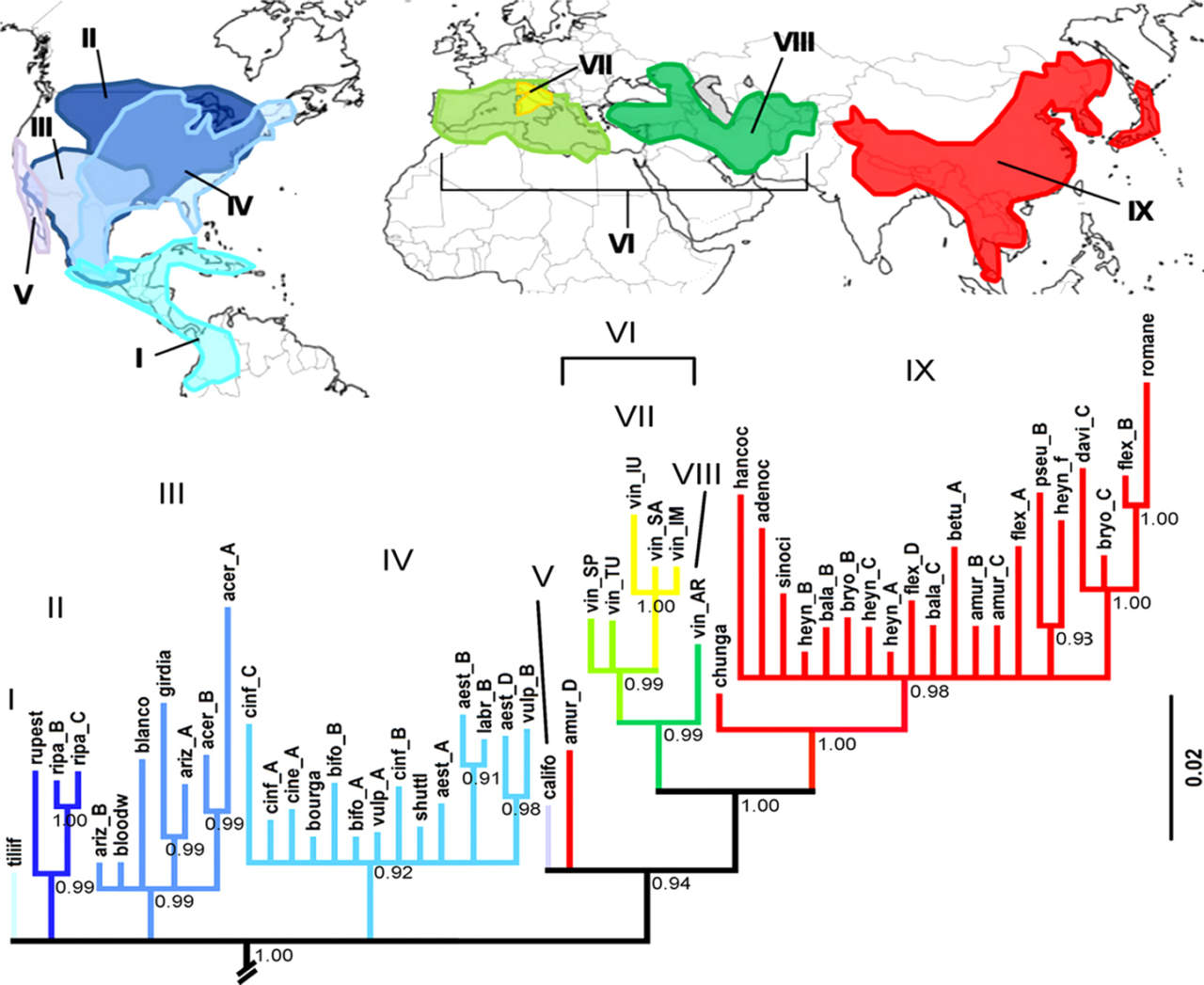
«Prima dell’avvento della tassonomia molecolare – ha detto - si è ad esempio dibattuto molto se la vite si fosse formata nel continente americano o asiatico, considerato che entrambi i continenti presentano un notevole pool genetico rappresentato da una trentina di specie ciascuno».
Un’origine oggi svelata da modelli filogenetici che la posizionano in Nord America. «Tra i molti lavori sperimentali su questo tema, facendo in particolare riferimento a quello del gruppo di Milano (Zecca e coll.), emerge con chiarezza che le specie americane rappresentino il pool ancestrale, poi frammentatosi durante il Miocene, con la successiva differenziazione delle specie asiatiche nel contesto del cambiamento tettonico e climatico del Pliocene-Quaternario».
V.vinifera è dunque più vicina alle specie asiatiche come V.amuriensis piuttosto che a quelle americane.
«L’origine delle specie asiatiche da quelle americane - ha chiarito Testolin - è facile da comprendere se, anzichè mettere l‘Europa al centro del mondo, lo inquadriamo dalla prospettiva polare che mette l’America del Nord vicino all’Asia».
«I due continenti hanno avuto scambi continui se consideriamo che le glaciazioni hanno spesso portato all’emersione di “ponti” terrestri tra i due continenti (oggi distanti solo 80 km con un fondale di profondità di 30 – 50 metri)». In definitiva il genere Vitis potrebbe aver fatto lo stesso percorso dell’H.sapiens, che è passato dall’Asia alle Americhe attraverso lo stretto di Bering, ma al contrario.
Le specie americane sono dunque quelle ancestrali, le asiatiche sono derivate da quelle americane e la vite europea avrebbe avuto origine dalle viti asiatiche, che nel frattempo avevano colonizzato l’Asia centrale e il Caucaso.
L’identità sessuale
Sono 370 le varietà resistenti registrate e dietro l’angolo, in attesa che si realizzino le promesse delle Nbt
Un altro rompicapo che si sta componendo è quello relativo all’”identità sessuale” di V. vinifera, così differente da quello delle altre specie di questo genere perché è l’unica caratterizzata da ermafroditismo (presenta nello stesso fiore sia gli organi maschili che femminili) e cleistogamia (fecondazione autogama che avviene “a fiore chiuso”).
«Dal punto di vista evolutivo l’ermafroditismo è lo stato ancestrale, ma la dioicia (sessi diversi su individui distinti) è un fenomeno molto diffuso nel regno vegetale. È infatti presente in oltre 15 mila specie distribuite in 175 famiglie e quasi mille generi». La dioicia favorisce la diversità genetica e l’adattamento evolutivo e la sua diffusione testimonia che il fenomeno è comparso in maniera indipendente centinaia di volte durante l’evoluzione.
Le viti selvatiche sono tutte dioiche, con piante maschili e femminili. Come è avvenuto il passaggio dalla diocia alla cleistogamia in V.vinifera?
«Il fatto interessante - ha messo in evidenza Testolin - è che mentre il fenomeno dal punto di vista fenotipico è piuttosto costante (fiori maschili caratterizzati da stami regolari e ovario atrofizzato e fiori femminili con polline sterile e ovario ben sviluppato), i meccanismi genetici coinvolti sono molti nelle diverse specie».
Nella vite la dioicia è controllata da 2 geni:
- maschio sterilità (gene Inp1): il polline non forma i pori per la germinazione e si hanno fiori fisiologicamente femminili;
- aborto dell’ovario (gene VviPLATZy). Nel fiore diventato maschile, l’ovario non si sviluppa per via di un gene (il VviPLATZy appunto), che riduce la produzione di citochinine e impedisce il normale sviluppo dell’ovario.
«Il ritorno dalla dioicia all’ermafroditismo è semplice perché richiede solo 1 o 2 mutazioni, mentre il controllo della cleistogamia non è noto ma potrebbe essere associato al ritorno all’ermafroditismo nella vite coltivata. Il fatto che siano presenti solo in V. vinifera testimonia che sono fenomeni recenti».
L’addomesticamento in Oriente
La storia della vite coltivata è scritta all’interno dei suoi geni e alcuni recenti lavori sperimentali (Laucou et al 2018 PLOS One; This et al 2006 Trends Genetics; Wales et al 2016 J Archeo Science; Zhou et al 2017 PNAS Liang et al 2019 Nature Comm.) hanno cercato di decodificare le tappe dell’addomesticamento in Eurasia.
«Ci sono 7 centri primari e secondari di addomesticamento ma durante questo tragitto millenario tra Caucaso e mediterraneo orientale, Medio Oriente, Balcani, penisola italiana, Europa occidentale e centrale, penisola iberica, Russia e Ucraine e Maghreb si è registrato un forte mescolamento delle varietà e introgressioni di geni da V. sylvestris».
L’analisi dei cloroplasti ha riscontrato che i cpDNA A,B,C sono comuni tra i resti archeologi e le varietà moderne, tuttavia alcuni lavori recenti stanno facendo luce sul tema dell’origine delle varietà moderne (Myles et al 2011 PNAS; This et al 2006 Trends Genet.; D’Onofrio et al 2021 Frontiers PS). Riguardo a questo tema è importante sottolineare che ci sono tre problemi che rendono difficili le ricostruzioni.
Testolin ha infatti evidenziato a Udine che:
- manca la letteratura antecedente al 1200 dC;
- fino al 1800 il vino era più importante del vitigno;
- i risultati riflettono il campionamento delle accessioni utilizzate per le analisi.

«L’ultimo è il limite maggiore: i risultati della letteratura scientifica riflettono il campionamento delle accessioni utilizzate per le analisi e a mancare è sempre il materiale caucasico che, con grande difficoltà e molto merito, hanno iniziato a raccogliere da alcuni anni i colleghi di Milano (Attilio Scienza, Osvaldo Failla, Laura Rustioni) assieme a quelli georgiani (David Maghradze)».
Il superamento di questi limiti sta consentendo di mettere in evidenza relazioni di parentela un tempo impensabili. «I funding clones, cioè le varietà capostipiti, che hanno dato origine a quelle che coltiviamo, non sono le varietà più note, ma germoplasma spesso trascurato e considerato geneticamente non interessante». Anche oggi i genetisti, nel pianificare gli incroci, a volte utilizzano queste informazioni sui funding clones. «Si sfruttano le informazioni storiche per evitare di eseguire costosi studi di attitudine combinativa e progeny test».
È così che, a titolo di esempio, il Gouais, vitigno che l’amministrazione francese ha provato più volte ad eliminare, è nel pedigree di 250 varietà (spesso insieme al Pinot). «In termini zootecnici si direbbe che è un cattivo produttore ma un buon “razzatore”».
Qualcosa di simile è capitato per il germoplasma nazionale. Il lavoro di D’Onofrio (ne abbiamo parlato nell’editoriale di VVQ 4) fa emergere infatti funding clones sottovalutati a cui dovremmo prestare più attenzione: Heunisch weiss, Malvasie, Achladi, Mantonico, Visparola, Bombino bianco, Garganega.
La libera ricombinazione di questo materiale genetico ha dato origine a tutte e varietà adattate alle diverse condizioni pedoclimatiche del nostro territorio. «Ho visitato di recente due produttori vitivinicoli etnei: è incredibile l’adattamento a climi e suoli così particolari che mostrano varietà come Nerello Mascalese e Carricante».
Breeding per una viticoltura sostenibile
L’arrivo di peronospora e oidio dalle Americhe ha costituito un evento drammatico per la viticoltura europea, ma l’aspetto positivo è che ha spinto a sviluppare programmi di incrocio e selezione con l’obiettivo della resistenza. Oggi sono attivi diversi centri che applicano tecniche di breeding, spesso assistito da marcatori molecolari, in 25 Paesi, che hanno sviluppato 370 varietà resistenti già registrate per la coltivazione. «Selezioni con caratteri del tutto simili a V. vinifera».
Dietro l’angolo c’è però già il miglioramento genetico di precisione promesso dalle New breeding technologies. La cisgenesi realizza una trasformazione genetica in cui geni e promotori provengono dalla stessa specie o da specie sessualmente compatibili. Le attività con questa tecnica sono limitate. «Ci lavorano sostanzialmente solo il Crea -VE di Conegliano, la Fondazione Edmund Mach e l’IGA di Udine lavorando su un gene di resistenza a peronospora (rpv 3-1) e uno di resistenza ad oidio del gruppo MLO».
«La frontiera più promettente è però quella del genome editing tramite Crispr/Cas9 (vedi alcuni esempi di lavori su vite nella tabella). Una tecnica tanto innovativa e facile da realizzare da mettere paura anche alle sue inventrici Emmanuelle Charpentier e Jennifer Doudna, entrambe premio Nobel per la chimica nel 2020».
| Genome editing, le applicazioni in corso su vite | |||||
| Ente | Gene | Funzione | Metodo | Status | Riferimento |
| CNR-Torino | [reporter] | - | CRISP-Cas9 | In progress | Perrone 2020 com pers |
| FEM | MLO-7 | Resistenza a oidio | CRISP-Cas9 | In progress | Malnoy et al 2016 |
| Dalla Costa et al 2019 | |||||
| CREA-VE | mlo06/mlo07 | Resistenza a oidio | CRISP-Cas9 | In progress | Kusch & Panstruga 2017 |
| CREA-VE | Vvagl11 | Apirenia | CRISP-Cas9 e Cas12 | In progress | Velasco 2021 com pers |
| IGA /VCR | [none] | - | CRISP-Cas9 | In progress | Testolin 2021 com pers |
| UNIVR/EDIVITE | VvAGL11/ MADSbox5 | Apirenia | CRISP-Cas9 | In progress | Zenoni 2020 VVQ |
La tradizione brandita come una clava
«Ciò che caratterizza questi protocolli, soprattutto quelli in cui la trasformazione riguarda la modifica di una o poche basi nella sequenza del DNA originale, è che non resta traccia di quanto avvenuto, a parte ovviamente le basi azotate modificate». «Verrebbe da dire a chi ha tanto criticato gli Ogm invocando il principio di precauzione che, se non si voleva che restasse traccia della trasformazione, bastava aspettare e lasciare lavorare i ricercatori in pace».
Si può sempre recuperare: «rompere gli ormeggi e navigare per l’aperto mare», usando un’espressione cara a Testolin. Anche perché gli obiettivi che impegnano i genetisti su vite sono importanti e impegnativi (vedi riquadro in alto).
In definitiva il rispetto della tradizione è importante, ma non può essere impiegato come una clava contro l’innovazione. La vite infatti si evolve da milioni di anni e continuerà a farlo (senza contare su di noi) nonostante le pretese di immutabilità dei vitigni coltivati. Possiamo essere semplici spettatori di questa evoluzione oppure, se vogliamo continuare questo percorso comune con V. vinifera, non possiamo che puntare sulle migliori tecniche di breeding.








{kind=link}